Structural characterisation and biological activities of a unique type β-d-glucan obtained from Aureobasidium pullulans
Abstract
A β-D-glucan obtained from Aureobasidium pullulans (AP-FBG) exhibits various biological activities: it exhibits antitumour and antiosteoporotic effects and prevents food allergies. An unambiguous structural characterisation of AP-FBG is still awaited. The biological effects of β-D-glucan are known to depend on its primary structures, conformation, and molecular weight. Here, we elucidate the primary structure of AP-FBG by NMR spectroscopy, and evaluate its biological activities. Its structure was shown to comprise a mixture of a 1-3-β-D-glucan backbone with single 1-6-β-D-glucopyranosyl side-branching units every two residues (major structure) and a 1-3-β-D-glucan backbone with single 1-6-β-D-glucopyranosyl side-branching units every three residues (minor structure). Furthermore, this β-D-glucan exhibited immunostimulatory effects such as the accumulation of immune cells and priming effects against enterobacterium. To our knowledge, 1-3-β-glucans like AP-FBG with such a high number of 1-6-β-glucopyranosyl side branching have a unique structure; nevertheless, many 1-3-β-glucans were isolated from various sources, e.g. fungi, bacteria, and plants.
β-Glucan is a well-known biological response modifier (BRM) widely distributed in nature. A variety of β-glucans have been isolated from various sources, e.g. fungi, plants, and seaweeds. The physicochemical properties of β-glucans differ based on their primary structure, including linkage type; degree of branching (DB); degree of polymerisation (DP); conformation, e.g. triple helix, single helix, and random coil structures; and molecular weight [1–3]. Recent reports have highlighted a significant role of β-glucans in the treatment of cancer and infectious diseases in both modern medicine and traditional oriental therapies. It also plays an important role as a dietary substance because it lowers the plasma cholesterol level, enhances the hematopoietic response, and possesses antitumour and immunomodulating properties [4–6]. For instance, lentinan from Lentinus edodes [7] and sonifilan (SPG) from Schizophyllum commune [8] have been used clinically for cancer therapy in Japan.
The polymorphic fungus Aureobasidium pullulans (A. pullulans) has been the subject of intense academic and industrial interest [9, 10]. In particular, the β-glucan secreted by A. pullulans has received much attention. Numerous researchers reported that the β-glucan prepared by A. pullulans exhibits not only an antitumour effect [11], but also various biological activities. For example, it promotes the DNA synthesis of human peripheral blood mononuclear cells (PBMCs), thus, inducing the production of cytokines [12], and preventing food allergies [13]; it also exhibits antiosteoporotic effect in ovariectomized osteoporotic mice [14]. However, molecular information regarding these effects, including the precise structure and underlying mechanism(s), is not completely understood.
Host molecules that serve as β-glucan receptors have recently been reported, e.g. complement receptor type 3 (CR3) [15] and lactosylceramide (LacCer) [16]. More recently, dectin-1 has also been described as a pivotal receptor for 1-3-β-glucan [17, 18]. It is important to know the relationship between the structure and receptor-binding ability of β-glucans [19]. Although it is well established that the biological effects of β-glucan depend on its primary structure, conformation, and molecular weight [1, 2], the precise structure of a β-glucan extracted from the black yeast A. pullulans, which is termed A. pullulans-fermented β-glucan (AP-FBG), is still unclear. Therefore, characterisation of the structure by nuclear magnetic resonance (NMR) spectroscopy of AP-FBG is important not only for promoting its clinical usage in immunotherapy, but also for understanding the mechanisms underlying its biological effects.
In the present study, we report the primary structure of AP-FBG and completely assign all its protons and carbons by 1D- and 2D-NMR spectroscopy, including correlation spectroscopy (COSY), total correlation spectroscopy (TOCSY), nuclear Overhauser effect spectroscopy (NOESY), heteronuclear single quantum coherence spectroscopy (HSQC), HSQC-TOCSY, heteronuclear multiple bond coherence spectroscopy (HMBC), and heteronuclear 2-bond correlation spectroscopy (H2BC) experiments, and also report its biological activities.
2 Materials and methods
2.1 Materials
D2O (deuteration degree min. 99.96%) and Me2SO-d6 (99.96%) were purchased from Merck. Grifolan was prepared as described previously [20]. β-Glucan from Candida albicans (CSBG) was prepared as described previously [21]. Pullulan, a β-glucan extracted from the Shiitake mushroom L. edodes, and soluble starch were purchased from Wako Pure Chemical Industries, Ltd. (Osaka, Japan). Cellotetraose and laminaritetraose were obtained from Sikagaku Corporation (Tokyo, Japan). Saline (Otsuka Pharmaceutical Co., Tokyo, Japan), phosphate-buffered saline (PBS) and RPMI-1640 (Nissui Seiyaku Co., Tokyo Japan), foetal calf serum (FCS; Daiichi Kagaku Yakuhin Co., Tokyo, Japan), Diff-Quik stain (Kokusai Shiyaku K.K., Kobe, Japan), penicillin-streptomycin (P4333; SIGMA-ALDRICH, St. Louis, MO), indigestive dextrin (Matsutani Chemical Industry Co., Ltd., Osaka, Japan), and guar gum hydrolysate (Taiyo Kagaku Co., Ltd., Tokyo, Japan) were also used in this study. For the cytokine measurement, enzyme-linked immunosorbent assay (ELISA) kits (Becton, Dickinson and Company, USA) were used.
2.2 Mice
In this study, we used 5-week-old male ICR mice purchased from CLEA Japan, Inc. (Tokyo, Japan). The animal experiments in the present study were conducted in accordance with the animal experimentation guidelines of ADEKA Corporation and Teikyo Univ., on the basis of the established guidelines by the Ministry of Education, Culture, Sports, Science and Technology.
2.3 Yeasts and culture conditions
The fugal strain A. pullulans ADK-34 (FERM BP-8391), isolated by ADEKA Corporation, was maintained on YM agar (1% D-glucose, 0.5% peptone, 0.3% yeast extract, 0.3% malt extract, 1.5% agar, pH 6.0). The A. pullulans ADK-34 strain was cultured, and the procedure used was as follows: 3 l of a SYE medium (3% sucrose, 0.5% yeast extract, 0.025% K2HPO4, 0.0125% MgSO4, 0.00025% FeSO4, pH 6.5) was added to a fermenter, and inoculated yeast precultured in the YM medium for 2 days at 26°C was cultured for 3 days at 26°C with air supplied at a rate of 1.5 l/h, rotation at 300 rpm, and pH maintained at 5.0.
2.4 Preparation of AP-FBG
Following the culture described above, the obtained culture medium was heated at 85°C for 30 min to kill the yeast. After centrifugation for 15 min at 5,000 rpm, the supernatant was carefully collected. Then, a 2-equivalent volume of ethanol was added and the precipitate was collected. After the precipitate was dissolved in distilled water, a 2-equivalent volume of ethanol was added and the precipitate was again collected. This procedure was repeated three times. The final precipitate was dialysed against excessive distilled water. The dialysed solution was lyophilised, and AP-FBG was obtained (yield, 12.7 g).
2.5 NMR spectroscopy
Exchangeable protons were removed by suspending the AP-FBG in D2O and lyophilising it. This exchange process was repeated 3 times. The method of Kim et al. [3] was used to record all the spectra in a mixed solvent of Me2SO-d6/D2O (6:1; 20 mg/ml) at 70°C, on a Bruker Avance 500 spectrometer equipped with a TXI-xyz 3 gradient probe for detecting 1H or a BBO-z gradient probe for detecting 13C. We used the internal Me2SO signal (δH = 2.53 ppm) for 1H and internal Me2SO (δC = 39.5 ppm) for 13C as references, and gave the chemical shifts in ppm. The Bruker standard pulse sequence was used to perform the 1D-1H experiment with a spectral width of 3,180 Hz in 64K complex data points. The relaxation delay used was 5T1 in order to calculate accurate signal integrations. Prior to Fourier transformation, zero filling was used 4 times, and noise was reduced with the TRAF function. The Bruker standard pulse sequence was used to perform the 1D-13C experiment, with a spectral width of 30,581 Hz in 64K complex data points. Prior to Fourier transformation, zero filling was used four times, and noise was reduced by exponential multiplication. The Bruker standard pulse sequence was used to conduct two-dimensional 1H COSY with 768 increments of 4,096 data points with 16 scans per t1 increment. The spectral width was 2,741 Hz in each dimension. Two-dimensional TOCSY was carried out with a TOCSY spinlock mixing time of 30–150 ms; the pulse sequence of Griesinger et al. was used to suppress rotating-frame Overhauser effect (ROE) signals [22]. The spectral width was 3004 Hz in each dimension, and 512 increments of 4,096 data points with 16 scans per t1 increment were recorded. Two-dimensional NOESY was conducted with a mixing time of 200 ms, with the Bruker standard pulse sequence. The spectral width was 2,741 Hz in each dimension, and 512 increments of 4,096 data points with 32 scans per t1 increment were recorded. The Bruker standard pulse sequence was used to conduct two-dimensional 13C-edited HSQC with 512 increments of 2,048 data points with 32 scans per t1 increment. The spectral width was 2,741 Hz for t2 and 11,061 Hz for t1. The Bruker standard pulse sequence was used to conduct two-dimensional 1H, 13C-HMBC with 256 increments of 2,048 data points with 132 scans per t1 increment. The delay time for the evolution of long-range couplings was set to 50 ms (optimized for 10 Hz). The spectral width was 2,948 Hz for t2 and 11,061 Hz for t1. The Bruker standard pulse sequence was used to conduct two-dimensional 1H, 13C-HSQC-TOCSY (a TOCSY spinlock mixing time of 30–120 ms was used) with 512 increments of 2,048 data points with 64 scans per t1 increment. The spectral width was 2,741 Hz for t2 and 11,061 Hz for t1. The pulse sequence of Nyberg et al. [23–25] was used to conduct two-dimensional H2BC with 256 increments of 2,048 data points with 128 scans per t1 increment. The spectral width was 5,000 Hz for t2 and 26000 Hz for t1. Prior to Fourier transformation, all 2D experiments were zero filled to 2K and 2K in both dimensions. A square cosine-bell window function was applied in both dimensions, except in the COSY and H2BC experiments. For the COSY experiment, a sine-bell window function was applied in both dimensions. The H2BC experiment was treated with a cosine window function in t1 and a π/4-shifted sine in t2.
2.6 Preparation of peritoneal exudate cells (PECs) and counting of cell populations
The preparation and counting of PECs was performed according to a previous study by Morikawa et al. [26]. Saline of the samples was injected into the peritoneal cavity of mice and at appropriate times, PECs were harvested with 7 ml of PBS. The cells were then washed and suspended in 2 ml of PBS containing 10% heat-inactivated FCS; following this, they were fixed on a glass slide with methanol and stained with Diff-Quik stain. The populations of polymorphonuclear cells (PMNs), macrophages, and lymphocytes in the preparation obtained from each mouse were determined microscopically by scoring 150 ± 10 cells in three fields selected at random on the slide. The mean values for three mice were calculated. The total number of leukocytes was counted microscopically with Turk’s reagent, and its value was calculated in each mouse.
2.7 PEC cultures and measurement of cytokines
AP-FBG (1 mg) was injected into mice. After 96 h, PECs were harvested and suspended in RPMI-1640 containing 10% FCS, 1% glutamate, and 100 units of penicillin-streptomycin, and the cell number was adjusted to 1 × 106 cells/ml. The cell suspension (200 µl) was plated on 96-well multiplates with 50 µg/ml of heat-killed cells [Enterococcus faecalis (E. faecalis) strain EF-2001] prepared by the Lab. of Medical Life Chemistry, Teikyo Univ., and then cultured for 2 or 24 h in 5% CO2. The cytokine content of both culture media was measured by ELISA. The 2 h-culture was used to detect tumour necrosis factor (TNF)-α, while the 24 h-culture was used to assess the remaining cytokines.
2.8 ELISA for detecting anti-β-glucan antibody
A 96-well Nunc plate was coated with a glucan preparation in 0.1 M carbonate buffer (pH 9.6) by overnight incubation at 4°C. The plate was washed with PBS containing 0.05% Tween 20 (PBST) and blocked with 1% bovine serum albumin (BSA) (BPBST) at 37°C for 60 min. After washing the plate, we incubated it with human immunoglobulin (polyglobinN) at 37°C for 60 min. The plate was then washed with PBST and treated with peroxidase-conjugated anti-human IgG + M + A antibody in BPBST, and developed with a tetramethylbenzidine (TMB) substrate system. Colour development was arrested with 1 M phosphoric acid, and the optical density was measured at 450 nm.
3 Results
3.1 Structural characterisation of AP-FBG by NMR spectroscopy
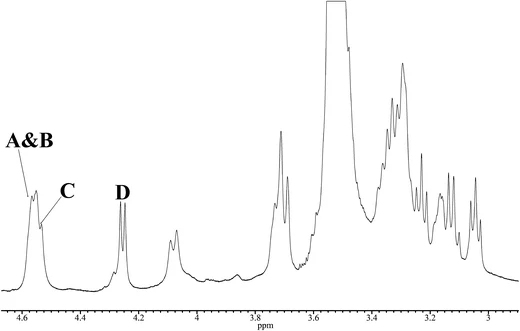
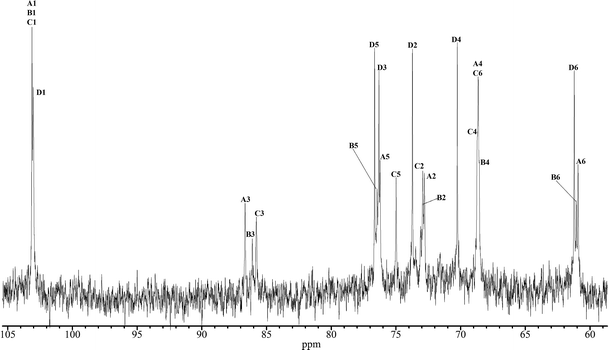
To characterise the structure of a β-glucan from A. pullulans used in this study (AP-FBG), we studied the structure of AP-FBG by NMR spectroscopy. A 1D-1H NMR spectrum of a solution of Me2SO-d6/D2O (6:1) at 70°C is shown in Fig. 1. The anomeric region (δH 4.2–4.7 ppm) contained four signals, three of which heavily overlapped (δH 4.55 ppm). The other peak was of a well-resolved doublet resonance (δH 4.255 ppm). The overlap of the anomeric doublets was confirmed by additional NMR experiments, including a COSY experiment. The 4 sugar residues in AP-FBG were arbitrarily labelled A, B, C, and D, as described in Fig. 1. On the basis of their observed chemical shifts, 3JH1, H2 and 1JH1, C1 (Table 1), all the residues were assigned as β-hexapyranosyl residues. The 1D-13C NMR spectra in Fig. 2 show 2 signals in the anomeric region (δC 95–110 ppm); they were assigned to residues A, B, and C (δC 103.03 ppm) and residue D (δC 103.12 ppm), which were confirmed by cross-peaks in the 1H, 13C-HSQC spectrum.
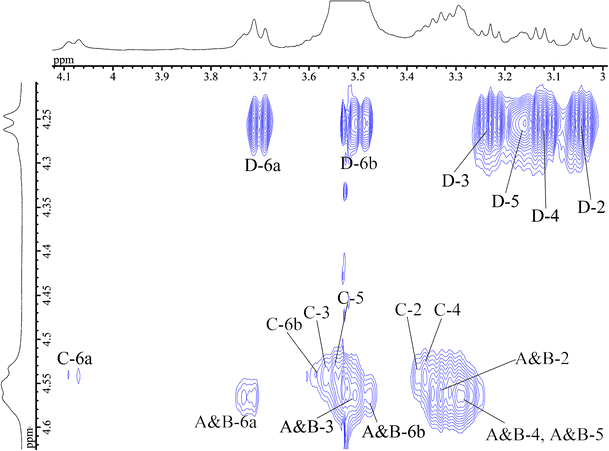
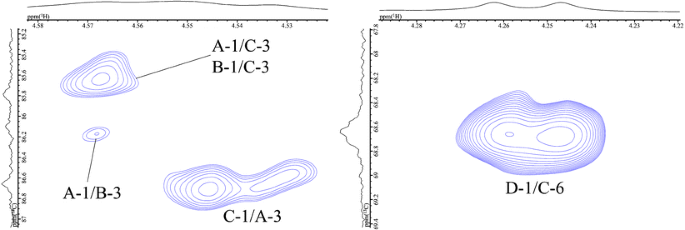
Next, we attempted to assign all 1H resonances in the 1H NMR spectrum based on the COSY and TOCSY experiments. In the 2D-TOCSY spectrum (Fig. 3), a complete series of cross-peaks was observed between A H-1 and A H-2, 3, 4, 5, 6a, and 6b, as well as between B, C, and D H-1 and B, C, D H-2, 3, 4, 5, 6a, and 6b. However, assignment of some of their protons (i.e., between A H-2 and H-4; and A H-4 and H-5; residues A H-2 to H-6 and B H-2 to H-6) were ambiguous, due to extensive overlap with the neighbouring protons. We further studied these ambiguously assigned protons by using the overlaid spectra of 13C-edited HSQC and H2BC, in order to separate the neighbouring protons in the 13C dimension. That is, the overlaid HSQC and H2BC spectra traced out the entire intra-ring assignment, as reported previously [23, 27]. Thus, all the protons in AP-FBG were assigned unambiguously.
Fig. 3
Anomeric region of the 2D-TOCSY spectrum of AP-FBG at 70°C. The 2D-TOCSY experiment was performed with a spinlock mixing time of 150 ms
In addition, the combination of the 13C-edited HSQC and HSQC-TOCSY spectra with various mixing times (30 to 120 ms) for TOCSY spinlock and the H2BC experiment allowed the complete assignment of the 13C spectrum. Table 2 summarizes the 1H and 13C NMR spectral assignments of AP-FBG. These assignments are based on the COSY, TOCSY, HSQC, HSQC-TOCSY, and H2BC spectra.
Table 2 Chemical shifts (ppm) of 1H and 13C NMR signals for AP-FBG
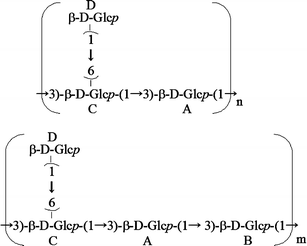
In the spectrum of AP-FBG, five cross-peaks (i.e. between A H-1 and C H-3, between A H-1 and B H-3, between B H-1 and C H-3, between C H-1 and A H-3, and between D H-1 and C H-6), which are shown in the 1H, 13C-HMBC spectrum (Fig. 4), were assigned to glycosidic linkages, as indicated by A(1→3)C, A(1→3)B, B(1→3)C, C(1→3)A, and D(1→6)C, respectively. These results were also confirmed by the NOESY experiment (data not shown). The ratio of the main chain (residues A, B, and C) to the side chain (residue D) was approximately 2:13 when calculated based on the accurate integrities of anomeric protons. In addition, the signal intensity and integrity of residue B were less than those of the other residues of the main chain (A and C). Therefore, these results strongly suggested that the primary structure of AP-FBG contains 1-3-β-glucan and 1-6-β-glucan units, as described in Fig. 5. As shown in this figure, AP-FBG analysed in this study has a mixed structure comprising a 1-3-β-D-glucan backbone with single 1-6-β-D-glucopyranosyl side-branching units every two residues as the major structure and a 1-3-β-D-glucan backbone with single 1-6-β-D-glucopyranosyl side-branching units every three residues as the minor structure. In addition, the ratio of major and minor units was approximately 7:3 when calculated based on the signal integrities of C-3 position in the 13C NMR spectrum.
3.2 Effect of AP-FBG on the cell number and population of PECs
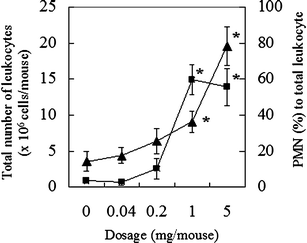
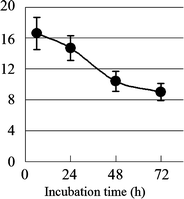
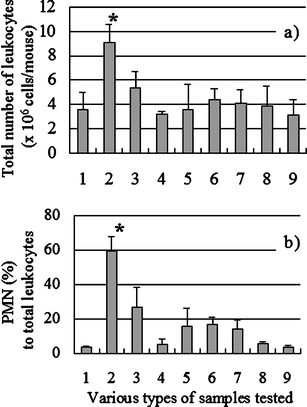
The primary structure of AP-FBG was clarified by NMR analysis in this study; therefore, the biological activities of AP-FBG were investigated. First, we investigated the change in the population of PMNs and total leukocytes in the peritoneal cavity of mice intraperitoneally administered with AP-FBG. As shown in Fig. 6, following AP-FBG injection, the number of total leukocytes and population of PMNs increased in a dose-dependent manner. Next, changes in the populations of PMNs, macrophages, lymphocytes, and total leukocytes were examined and shown in Fig. 7. Total leukocytes reached a peak after 6 h, and then decreased from 6 to 72 h. In detail, the peak number of PMNs (>60%) was observed at 6 h. It decreased from 6 to 24 h (<10%), while the macrophage count increased progressively from 6 to 72 h (Table 3). Furthermore, we conducted a comparison study between AP-FBG and other various water-soluble sugars by counting the ratio of PMNs to the total number of leukocytes. As shown in Fig. 8, AP-FBG administration resulted in an increase in the number of total leukocytes (Fig. 8a), and the PMN population (Fig. 8b) formed 60% of the PECs. This was statistically significantly different from that of all the other samples, including the β-glucan extracted from Shiitake mushrooms (L. edodes).
Fig. 6
Effects of AP-FBG doses ranging from 0 to 5 mg, on the total number of leukocytes and percentage of PMNs in the PECs. AP-FBG at 0, 0.04, 0.2, 1, and 5 mg in saline was intraperitoneally injected into each group of three mice, and PECs were harvested at 6 h. Then, the total number of leukocytes was counted, and the degree of PMN accumulation was calculated by microscopic observation [triangle, total leukocyte cell number (×106 cells/mouse); square, PMNs (%) to total number of leukocytes]. The results indicate the mean ± SD of 12 samples from three mice. *Statistically significantly different from the 0-mg group at p < 0.05
Time course of the changes in total leukocytes as a result of the AP-FBG injection. AP-FBG (5 mg) was injected intraperitoneally into 12 mice, and PECs were harvested at 6, 24, 48, and 72 h; PECs were harvested from three mice each time. Then, the total leukocytes were counted by microscopic observation
Comparison of various sugar samples with AP-FBG, with regard to the degree of PMN accumulation and total number of leukocytes in PECs. The various sugar samples and FBG at 1 mg were injected into groups of three mice each, and PECs were harvested at 6 h. Then, the total number of leukocytes (a) were counted and the proportion of PMNs in the total PECs was determined (b) by microscopic observation. No. 1 Negative control, No. 2 AP-FBG, No. 3 β-glucan extracted from the Shiitake mushroom Lentinus edodes, No. 4 pullulan, No. 5 soluble starch, No. 6 indigestive dextrin, No. 7 guar gum hydrolysate, No. 8 cellotetraose, No. 9 laminaritetraose. The results indicate the mean ± SD of the 12 samples from three mice. aStatistically significant difference between AP-FBG and all the other samples at p < 0.05
3.3 Priming effect of AP-FBG on the cytokine production induced by heat-killed E. faecalis
Next, we analysed the ability of AP-FBG to prime PECs against enterobacterium E. faecalis stimulation. As shown in Table 4, compared with the PECs derived from non-immunized mice, the production of TNF-α and interleukin (IL)-12 from the PECs of AP-FBG-immunized mice was significantly enhanced in response to heat-killed E. faecalis.
Table 4 Cytokines induced from the PECs of mice injected with AP-FBG
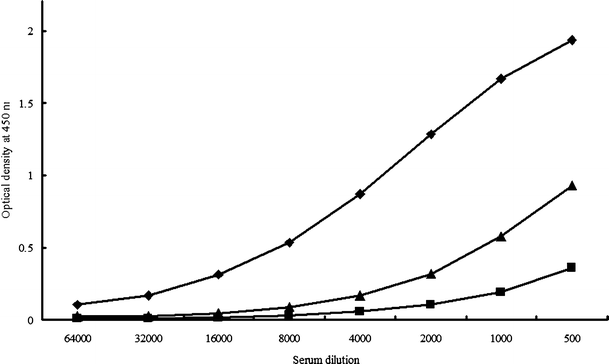
We previously reported the existence of anti-β-glucan antibodies in human [28] and mouse sera [29], although β-glucans are generally considered to be weak immunogens. Furthermore, these antibodies might be attributed to acquired immunity [30]. Thus, we investigated the reactivity of AP-FBG with human sera. We found that human sera exhibited high reactivity to AP-FBG; this reactivity was stronger than that to grifolan, which comprises a 1-3-β-D-glucan backbone with single 1-6-β-D-glucopyranosyl side-branching units every 3 residues, although it was weaker than that to CSBG, which comprises a 1-3-β-D-glucan backbone with long 1-6-β-D-glucopyranosyl side-branching units (Fig. 9).
Fig. 9
Reactivity of human sera to AP-FBG. The reactivity of AP-FBG (triangle), CSBG (diamond) and grifolan (square)was tested by ELISA. A 96-well plate was coated with the glucan preparation (25 µg/ml) in 0.1 M carbonate buffer (pH 9.6) by overnight incubation at 4°C. The plate was washed with PBST and blocked with 1% BPBST at 37°C for 1 h. After washing the plate, we incubated it with human sera at 37°C for 1 h. The plate was then washed with PBST and treated with the antibody for anti-human IgG + M + A in BPBST, and developed with a TMB substrate system. Colour development was arrested with 1 M phosphoric acid, and the optical density was measured at 450 nm
In the present study, we unambiguously elucidated the primary structure of β-glucan isolated from the black yeast A. pullulans, as a mixed structure comprising a 1-3-β-D-glucan backbone with single 1-6-β-D-glucopyranosyl side-branching units every 2 residues as the major structure and a 1-3-β-D-glucan backbone with single 1-6-β-D-glucopyranosyl side-branching units every three residues as the minor structure (the ratio of major and minor units was approximately 7:3). We also indicated that this β-glucan has potent adjuvant effects, that is, it acts as a BRM, and that the antibody against this β-glucan is present in human sera. These findings strongly suggested the prospective use of this unique type of β-glucan as an immunostimulatory agent in medicine.
Since the structure of a β-glucan from A. pullulans is still unclear, we initiated the unambiguous assignment of all 1H (Fig. 1 and Table 1) and 13C (Fig. 2) signals of AP-FBG, and clearly elucidated a primary structure by employing 1D- and 2D-NMR spectroscopy (Figs. 3 and 4 and Table 2). Previous reports by other groups on the structure of a β-glucan from another strain of A. pullulans used in this study have been insufficient, because these structural analyses were based on methylation and incomplete NMR analyses, and they only led to the elucidation of the tentative structure [13, 31]. The result showed AP-FBG to be composed of a mixture of two units: a 1-3-β-D-glucan backbone with single 1-6-β-D-glucopyranosyl side-branching units every two residues (major structure) and a 1-3-β-D-glucan backbone with single 1-6-β-D-glucopyranosyl side-branching units every three residues (minor structure) (the ratio of major and minor units was approximately 7:3).
The primary structure of AP-FBG was clarified by NMR analysis in this study; therefore, we examined its biological activities. AP-FBG administration in mice induced an increase in the number of total leukocytes and population of PMNs in a dose- and kinetics-dependent manner (Figs. 6 and 7 and Table 3). Morikawa reported [26] that the potential to induce PMNs and activate macrophages are essential qualities for BRMs, which can be classified into several types on the basis of the extent of their PMN response. Compared with curdlan (1-3-β-D-glucan), lentinan (1-3-β-D-glucan backbone with single 1-6-β-D-glucopyranosyl side-branching units every five residues), which has been used as an antitumour BRM, induced an increase and, consequently, a more rapid decrease in the number of PMNs. Simultaneously, the number of macrophages increased progressively; it therefore seems that the response to AP-FBG was categorized as the declining type (lentinan type). These results suggested that the responses to AP-FBG resembled the responses to lentinan rather than those to curdlan (data not shown), and we assumed that AP-FBG is a BRM classified as the lentinan type. These effects were more potent than that of all the other samples, including the β-glucan extracted from Shiitake mushrooms (L. edodes; Fig. 8). AP-FBG was also observed to possess potent adjuvant action (Table 4). These facts suggested that AP-FBG has superior potency as a BRM, because AP-FBG strongly induced a PMN accumulation response and activation of macrophages for the enhancement of cytokine production. In addition, since the anti-AP-FBG antibody was present in human sera (Fig. 9), innate as well as acquired immunity against β-glucans has been established in humans. This fact indicated that acquired as well as innate immunity might play a pivotal role in the in vivo biological response to β-glucan, i.e. AP-FBG.
In the present study, we examined the accumulation of leukocytes in PECs by administration of AP-FBG and the activities of the resulting PECs from the viewpoint of acute inflammatory response, and we found significantly strong activity against AP-FBG as compared to that against other glycan preparations. β-Glucan has been extensively investigated for over 30 years, and large amounts of data on various parameters have been accumulated. A tentative conclusion from these studies is the difficulty to evaluate the relative activities of glucan from one or few experimental evidences. The activities of β-glucan are affected at least by molecular weight, conformation, solubility and branching structure. β-Glucan activity also depends on the species and strains of the tested host. Thus, it might not be true that the best glucan always shows the strongest activity in all the assay protocols tested. Based on these facts, it is too early to derive any conclusion on the relative activity of AP-FBG compared to that of other glucan preparations. The experiment performed in this study revealed that AP-FBG could induce acute inflammatory response and priming effect to E. faecalis-induced cytokine synthesis. These data also strongly suggest that AP-FBG induced pattern recognition receptors (PRRs) on leukocytes for binding to pathogen-associated molecular patterns (PAMPs), especially receptors for E. faecalis, such as toll-like receptors (TLRs). Studies on the molecular mechanism(s) of the response of PRRs to AP-FBG might be valuable.
In conclusion, we revealed that the primary structure of β-glucan from A. pullulans comprised a mixture of a 1-3-β-D-glucan backbone with single 1-6-β-D-glucopyranosyl side-branching units every two residues as the major unit and a 1-3-β-D-glucan backbone with single 1-6-β-D-glucopyranosyl side-branching units every three residues as the minor unit (the ratio of major and minor units was approximately 7:3). Moreover, its immunostimulatory effects were also revealed. To our knowledge, the present data is the first to report the precise structural characterisation of β-glucan prepared from A. pullulans, by NMR spectroscopy. Few literatures have reported the highly branched structure of 1-3-β-glucans, e.g. SSG from the fungus Sclerotinia sclerotiorum [32] and OL-2 from the fungus Omphalia lapidescens [33]. 1-3-β-Glucans like AP-FBG with such a high number of 1-6-β-glucopyranosyl side branching have a unique structure; nevertheless, numerous 1-3-β-glucans were isolated from various sources, e.g. fungi, bacteria, and plants. Our data described here also suggested that although many papers have reported the structure of β-glucans by means of only 13C-NMR analysis, the existence of three signals for the C-3 position in the 13C-NMR spectra does not always correspond to the side-branching units that are present at every three residues. This information regarding structure can be used to gain insight into the molecular mechanisms involved in β-glucan-induced immune responses such as β-glucan-receptor interaction, by NMR spectroscopy, as well as the ligand specificity of β-glucan receptors.